Metabolismus and cell biology of unsaturated fatty acids
Metabolism and cellbiology of mono- and polyunsaturated fatty acids
Studies in Stearoyl-CoA desaturase (SCD1) and ω6-fatty acid desaturase (FADS-2) deficient mouse models
The cis double bond system of mono- and poly-unsaturated fatty acids imposes additional enzymes for positioning the double bonds in intermediates suitable for the ß-oxidation-cyle. The key enzymes are the mitochondrial 3cis-2-trans enoyl-CoA isomerase (ECI), responsible for the enzymatic transformation of odd numbered cis double bond systems of polyenoic fatty acids, 2,4-dienoyl-CoA reductase and 3-hydroxy-acyl-CoA-Epimerase for even numbered cis double bound.ECI completed our understanding of the degradation of all fatty acids in the ß-oxidation cycle. The rat isomerase has been cloned in this laboratory for detailed mechanistic understanding. We have crystallized the recombinant form of the mouse isomerase. The eci - null allelic mouse has been generated, which provided new insight and a diagnostic tool in pediatrics to discover a new form of inborn error among the numerous unknown mitochondrial defects in human.
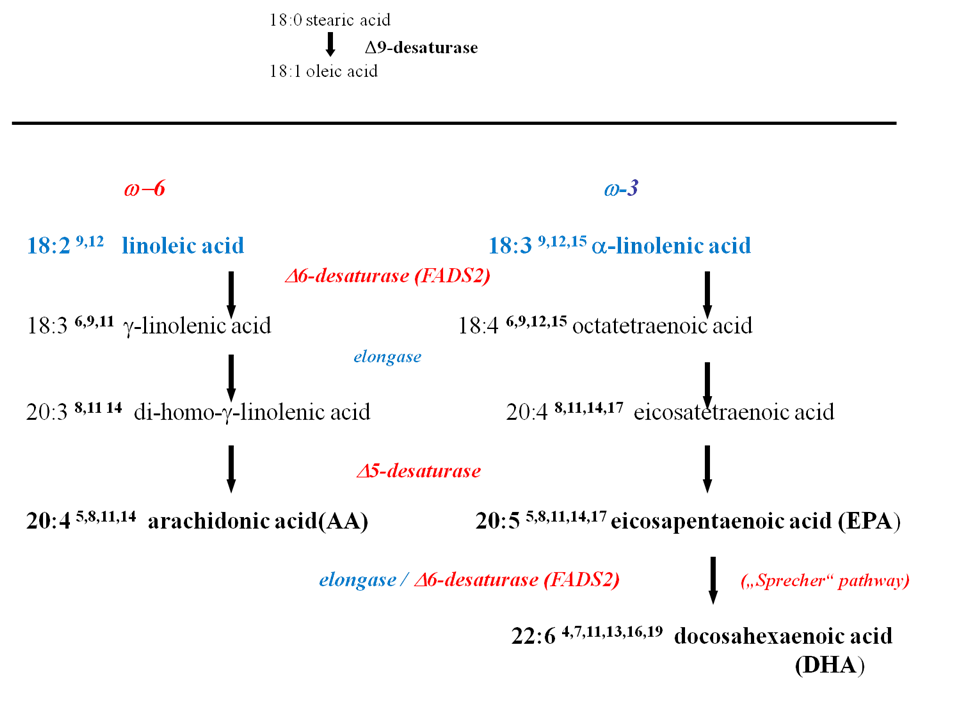
The essential fatty acids linoleic and alpha-linolenic acid are transformed to the eicosa- and docosapolyenoic fatty acid series in a cascade of desaturation and chain elongation reactions. We discovered that the chain elongation in this transformation cascade is catalyzed by the microsomal malonyl-CoA dependent fatty acid chain elongation complex and the initial desaturation is catalyzed by the ω6 fatty acid desaturase, both localized in the endoplasmatic reticulum (ER). De novo synthesized palmitoyl- and stearoyl- CoA are position- and stereospecifically desaturated by stearoyl-CoA desaturase (SCD-1), whereas the essential fatty acids are desaturated by ω6-fatty acid desaturase (FADS-2). To get insight into the role of mono- and poly-unsaturated fatty acids as membrane constituents, which provide pivotal biophysical properties of the membrane lipid bilayers and precursor functions of ω3- and ω6-PUFAs for biologically highly active signal molecules, we generated both the scd1-/- and fads2-/- knock out mouse mutants by gene targeting. We discovered the important role of SCD1 in transepidermal water barrier regulation, for energy and temperature homeostasis.
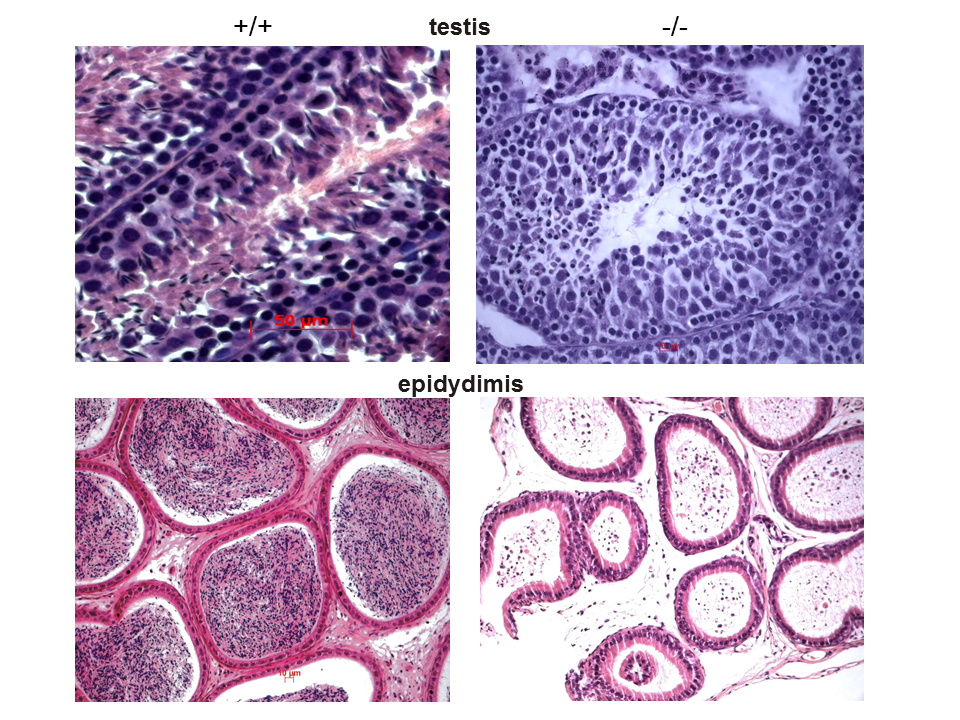
The lack of polyunsaturated fatty acids (PUFAs) in the fads2-/- mouse abolishes the precursor of eicosanoid synthesis in the cyclic (cyclooxygenases) and linear (lipoxygenase) pathways. FADS-2 deficiency disturbs the initial platelet aggregation step in the coagulation system. PUFA deficiency in phospholipids of the membrane lipid bilayer has a strong impact on membrane domain structures, e.g. leads to the disruption of intercellular adherens junction systems of the blood-testis barrier and causes male and female sterility. This barrier function defect together with male and female infertility can be restored specifically by dietary supplementation with ?3-docosahexaenoic acid. The many facets of the complex phenotype of the auxotrophic fads2-/- mutant are currently under investigation.
Biosynthetic pathways: from dietary ω3 (?18:3) and ω6 (18:2) essential fatty acids to ω3 and ω6 polyunsaturated fatty acids.
Sections of testis and epidiymis of wt and fads2-/- mice. Wt testis showed normal spermatogenesis and wt epididymis tubular system showed mature spermatozoa in the lumen of the seminiferous tubuli.
Induction of vascular injury and thrombosis in carotid artery (cross sections of carotid arteries). Thrombotic obliteration of the carotid artery of wt (+/+) mice and resistance to thrombosis in fads2-/- mice.